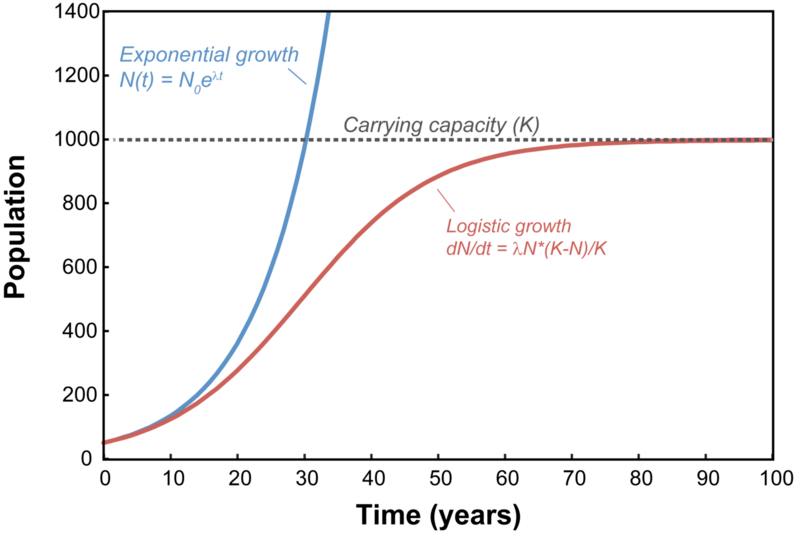
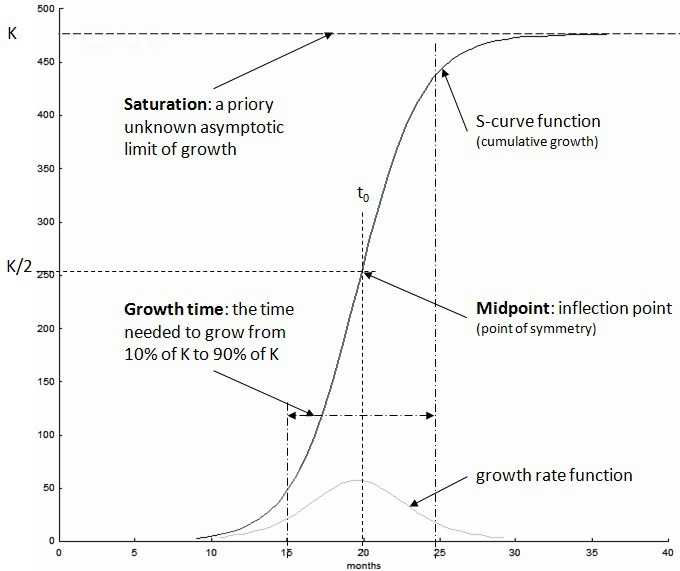
逻辑斯谛方程(Logistic Equation),又称逻辑斯谛增长模型,是一种描述在有限资源环境下种群规模增长的S形曲线数学模型。该模型由比利时数学家皮埃尔·弗朗索瓦·韦尔霍斯特(Pierre François Verhulst)于19世纪提出,如今已成为生态学、生物学、化学、人口学、经济学、地球科学、心理学、社会学、政治学、语言学、统计学、医学和机器学习等多个领域的核心模型之一。
重新发现与推广:韦尔霍斯特的工作在当时并未引起广泛关注。直到1920年,美国生物学家雷蒙德·珀尔(Raymond Pearl)和洛厄尔·里德(Lowell J. Reed)在研究果蝇种群增长和人口问题时独立重新发现了这一方程,并提出了现在广为流传的形式。他们的工作使得该方程在生物学界广为人知,因此该方程有时也被称为韦尔霍斯特-珀尔方程或“阻碍增长方程”[4]。
实验验证与理论拓展:1911年,安德森·格雷·麦肯德里克(Anderson Gray McKendrick)首次将逻辑斯谛方程用于描述肉汤中细菌的生长,并通过非线性参数估计进行了实验测试[5]。1925年,数学生物学家阿弗雷德·洛特卡(Alfred James Lotka)也独立推导出该方程,称其为“种群增长律”[6]。
混沌理论发展:随着20世纪系统科学和混沌理论的发展,该方程的离散形式——逻辑斯谛映射被深入研究。罗伯特·梅(Robert M. May)在1976年发表于《自然》杂志的论文《简单数学模型中的复杂动力学》中[7],系统阐述了这一简单非线性方程如何产生复杂的混沌行为,从而使该模型成为混沌研究的经典范例[8]。
Verhulst, P. F. (1838). Notice sur la loi que la population suit dans son accroissement. Correspondance mathématique et Physique, publiée par A. Quetelet, 10, 113-120.
Verhulst, P. F. (1845). Recherches mathématiques sur la loi d'accroissement de la population. Nouveaux Mémoires de l'Académie Royale des Sciences, des Lettres et des Beaux-Arts de Belgique, 18, 1-38.
Verhulst, P. F. (1847). Deuxième Mémoire sur la loi d'accroissement de la population. Nouveaux Mémoires de l'Académie Royale des Sciences, des Lettres et des Beaux-Arts de Belgique, 20, 1-32.
Pearl, R., & Reed, L. J. (1920). On the rate of growth of the population of the United States since 1790 and its mathematical representation. Proceedings of the National Academy of Sciences, 6(6), 275–288.
McKendrick, A. G., & Pai, M. K. (1912). XLV.—The Rate of Multiplication of Micro-organisms: A Mathematical Study. Proceedings of the Royal Society of Edinburgh, 31, 649-653.
Lotka, A. J. (1925). Elements of physical biology. Williams & Wilkins.
May, R. M. (1976). Simple mathematical models with very complicated dynamics. Nature, 261(5560), 459–467.
Strogatz, S. H. (2018). Nonlinear dynamics and chaos: with applications to physics, biology, chemistry, and engineering (2nd ed.). CRC Press.
Pianka, E. R. (1970). On r- and K-selection. The American Naturalist, 104(940), 592–597.
Begon, M., Townsend, C. R., & Harper, J. L. (2006). Ecology: from individuals to ecosystems (4th ed.). Blackwell Publishing.
Malthus, T. R. (1798). An essay on the principle of population. J. Johnson.
Kucharavy, D., & Roland, D. G. (2015). Application of logistic growth curve. Procedia Engineering, 131, 280-290.
Strogatz, S. H. (2018). Nonlinear dynamics and chaos: with applications to physics, biology, chemistry, and engineering (2nd ed.). CRC Press.
May, R. M. (1976). Simple mathematical models with very complicated dynamics. Nature, 261(5560), 459–467.
Feigenbaum, M. J. (1978). Quantitative universality for a class of nonlinear transformations. Journal of Statistical Physics, 19(1), 25–52.
Richards, F. J. (1959). A Flexible Growth Function for Empirical Use. Journal of Experimental Botany, 10(2), 290–301.
Tjørve, E., & Tjørve, K. M. C. (2010). A unified approach to the Richards-model family for use in growth analyses: why we need only two model forms. Journal of Theoretical Biology, 267(3), 417–425.
Hutchinson, G. E. (1948). Circular causal systems in ecology. Annals of the New York Academy of Sciences, 50(4), 221–246.
Fisher, R. A. (1937). The wave of advance of advantageous genes. Annals of Eugenics, 7(4), 355–369.
Gompertz, B. (1825). On the nature of the function expressive of the law of human mortality. Philosophical Transactions of the Royal Society of London, 115, 513–585.
West, G. B., Brown, J. H., & Enquist, B. J. (2001). A general model for ontogenetic growth. Nature, 413(6856), 628–631.
Tabatabai, M., Williams, D. K., & Bursac, Z. (2005). Hyperbolastic growth models: theory and application. Theoretical Biology and Medical Modelling, 30, 2–14.
Lotka, A. J. (1925). Elements of physical biology. Williams & Wilkins.
Volterra, V. (1926). Variazioni e fluttuazioni del numero d’individui in specie animali conviventi. Memorie della R. Accademia Nazionale dei Lincei, Serie VI, 2, 31–113.
Clark, C. W. (1990). Mathematical bioeconomics: the optimal management of renewable resources (2nd ed.). Wiley.
Cohen, J. E. (1995). How many people can the earth support? W. W. Norton & Company.
Lee, S. Y., Lei, B., & Mallick, B. (2020). Estimation of COVID-19 spread curves integrating global data and borrowing information. PLOS ONE, 15(7), e0236860.
Chowell, G., Hengartner, N. W., Castillo-Chavez, C., Fenimore, P. W., & Hyman, J. M. (2004). The basic reproductive number of Ebola and the effects of public health measures: the cases of Congo and Uganda. Journal of Theoretical Biology, 229(1), 119–126.
Roosa, K., Lee, Y., Luo, R., Kirpich, A., Rothenberg, R., Hyman, J. M., & Chowell, G. (2020). Real-time forecasts of the COVID-19 epidemic in China from February 5th to February 24th, 2020. Infectious Disease Modelling, 5, 256–263.
Simeoni, M., Magni, P., Cammia, C., De Nicolao, G., Croci, V., Pesenti, E., … & Rocchetti, M. (2004). Predictive pharmacokinetic-pharmacodynamic modeling of tumor growth kinetics in xenograft models after administration of anticancer agents. Cancer Research, 64(3), 1094–1101.
Epstein, I. R., & Pojman, J. A. (1998). An introduction to nonlinear chemical dynamics: oscillations, waves, patterns, and chaos. Oxford University Press.
Reif, F. (1965). Fundamentals of statistical and thermal physics. McGraw-Hill.
Stanley, H. E. (1971). Introduction to phase transitions and critical phenomena. Oxford University Press.
Bass, F. M. (1969). A new product growth for model consumer durables. Management Science, 15(5), 215–227.
Berkson, J. (1944). Application of the logistic function to bio-assay. Journal of the American Statistical Association, 39(227), 357–365.
Cox, D. R. (1958). Two further applications of a model for binary regression. Biometrika, 45(3/4), 562–565.
 发表于 2026-3-5 11:51:09
|
查看: 223|
回复: 0
发表于 2026-3-5 11:51:09
|
查看: 223|
回复: 0